
Response relationship between plant community diversity and altitude in Qilian Mountain National Nature Reserve, Gansu Province
-
摘要:目的 探求祁连山自然保护区植物群落多样性与海拔的响应关系及其驱动机制,为区域森林生态系统的保育和恢复提供一定的理论基础,同时也为区域生物多样性的保护利用及生态建设提供参考依据。方法 本研究在野外样方调查的基础上,综合运用除趋势典范对应分析法(Detrended canonical correspondence analysis, DCCA)、相关分析法及回归分析法,对甘肃祁连山自然保护区内(99°30′~100°53′E,38°12′~39°17′N,海拔1 500~3 700 m)植物群落多样性与海拔的响应关系进行了深入分析。结果 结果表明:① 不同层次植物群落丰富度指数、多样性指数在不同海拔表现出明显的差异,草本层和灌木层Patrick丰富度指数和Shannon-Wiener多样性指数随环境梯度和海拔梯度均表现出“单峰型”变化格局,除高海拔(2 950~3 700 m)外,在其余海拔梯度上,草本层丰富度指数和多样性指数均显著高于灌木层(P<0.01),且在中海拔(2 400~2 950 m)地带达到最高;② 植物群落各层次Pielou均匀度并未随环境梯度和海拔梯度表现出明显的差异,表明植物群落均匀度受环境响应较小,而主要受群落自身的结构类型和稳定程度的影响;③ 草本、灌木和乔木植被分布格局被环境因子解释部分分别占29.32%,23.24%和9.05%,被空间因子解释部分分别占15.21%,11.08%和7.52%,被空间和环境因子交叉作用的解释部分分别占11.04%、8.42%和5.31%,未被空间因子和环境因子解释部分分别占44.43%,57.26%和78.12%。结论 植物群落物种多样性除受海拔引起的水热梯度影响外,还受生物间相互作用、人类活动、发育程度等因素的影响,在未来生态系统的保护与管理中,在减少人类活动影响的同时,应增强应对气候变化的保护措施和力度。
-
关键词:
- 植物群落 /
- 海拔 /
- 祁连山 /
- 植物多样性 /
- 除趋势典范对应分析法
Abstract:Objectives To explore the response relationship between plant community diversity and altitude and its driving mechanism in Qilian Mountain Nature Reserve, and to identify the driving mechanisms behind the relationship. The findings will provide a theoretical basis for the conservation and restoration of the regional forest ecosystem, as well as a reference for the protection and utilization of regional biodiversity and ecological construction.Methods On the basis of field sample survey, Detrended canonical correspondence analysis (DCCA), correlation analysis and regression analysis were comprehensively used in this study. The relationship between plant community diversity and altitude in Qilian Mountain Nature Reserve (99°30′−100°53′E, 38°12′−39°17′N, altitude 1 500−3 700 m) was analyzed.Results The results showed that: 1) The richness index and diversity index of plant communities at different levels showed significant differences at different altitudes. The Patrick richness index and Shannon-Wiener diversity index of the herbaceous layer and the shrub layer showed a “unimodal” change pattern with environmental gradient and altitude gradients, except for high altitudes (2 950−3 700 m). In other elevations, the richness index and diversity index of the herbaceous layer were significantly higher than those of the shrub layer (P < 0.01) and reached the highest point in the middle elevation zone (2 400−2 950 m) zone. 2) The uniformity of Pielou did not significantly differ at each level of the plant community with the environmental and elevation gradients. This suggested that the uniformity of the plant community was less affected by the environmental response and was mainly influenced by the community’s structure type and stability. 3) The distribution pattern of herbaceous, shrub, and tree vegetation accounted for 29.32%, 23.24%, and 9.05% by environmental factors, 15.21%, 11.08%, and 7.52% by spatial factors, and 11.04%, 8.42% and 5.31% by the interaction of spatial and environmental factors, respectively. The proportion of unexplained by spatial factors and environmental factors was 44.43%, 57.26%, and 78.12%, respectively.Conclusions The species diversity of plant spieces within a community was influenced not only by the hydrothermal gradient caused by altitude, but also by biological interactions, human activities, development degree, and other factors. In the future protection and management of the ecosystem, measures must be taken to reduce the impact of human activities, and to strengthen efforts to cope with climate change. -
山地植被的生物多样性和空间分布格局能很好地反映植物利用环境资源的状况和种群在群落中的作用与地位,可以揭示植物个体之间相互依存、相互制约的复杂关系,对认识山地植物的生态适应性和维持植被群落稳定性机制具有重要意义[1]。山地植被的分布对土壤属性、温度和水分等外界因子具有很强的敏感性和直观性,其中,海拔作为综合性生态变量,包含了温度、水分、光照、土壤养分等因素,影响着山地植物的分布范围、群落动态及植被类型,被认为是区域植物群落空间分布格局和结构的决定性因子[2]。近年来,在全球气候变化的背景下,众多学者在不同区域、不同山脉对植物多样性沿海拔梯度的变化进行了大量研究[3]。例如,陈水飞等[4]在武夷山国家公园、王褚等[5]在天山北坡中部、徐翔等[6]在西双版纳研究中发现,植物群落物种多样性随海拔梯度分别呈单调下降、U型分布和单峰型分布的变化趋势,尚未形成统一结论[7]。因此,研究典型区域物种多样性与海拔的响应关系,不仅有助于研究植物多样性的维度格局,对深入了解植物多样性地理分布格局的形成机制也具有重要意义。
甘肃祁连山国家自然保护区地处青藏高原东北部,横跨河西走廊,属大陆性高寒半湿润山地气候[8],区域内物种多样性丰富,是河西走廊重要的生物物种资源库,对维持青藏高原生态平衡和西北干旱区流域生态安全具有重要的作用[9]。近年来,众多学者对祁连山国家自然保护区不同海拔梯度植物群落结构特征[10]以及土壤性状[11]、地形、气候、坡度[12]等地理环境因子对植物多样性的影响程度及驱动机理做了大量研究,其成果明确了祁连山自然保护区植物群落分布格局及其与环境因子的关系,为区域内天然林保护工程实施后森林资源恢复评价提供了参考。但以往研究多倾向于植物群落的结构特征及其与某几个环境因子的响应关系,尚未对不同海拔梯度植物群落多样性变化的驱动机制及变化机理进行深入探讨。
祁连山自然保护区北坡具有较大的海拔梯度,而海拔作为水热变化的综合性影响因子,因此,厘清研究区植物群落多样性的垂直变化效应及其驱动机制,对祁连山自然保护区精准制定植物多样性保护政策具有十分重要的意义。鉴于此,本研究以甘肃祁连山国家自然保护区内北坡植物群落为研究对象,在野外样方调查的基础上,采用除趋势典范对应分析法(Detrended canonical correspondence analysis, DCCA)和回归分析法,对研究区内不同植物群落物种多样性与海拔梯度的响应关系进行深入研究,以期揭示物种与生境之间的相互关系,为祁连山自然保护区森林生态系统的保育和恢复提供一定的理论基础,同时也为区域生物多样性的保护利用及生态建设提供参考依据。
1. 材料与方法
1.1 研究区概况
研究区位于甘肃祁连山国家自然保护区中段甘肃省张掖市境内,地处99°30′~100°53′E、38°12′~39°17′N之间(图1),受西风带的影响,具有高原和大陆性气候特征。不同海拔梯度,气候差异较大,降水主要集中在5—9月,年均气温和年均降水量分别为6 ℃和400 mm,无霜期一般为80~120 d。研究区内物种丰富,主要分布在1500~3700 m之间,具有明显的垂直梯带性[13],随着海拔梯度的升高,依次分布有超旱生荒漠灌丛[红砂(Reaumuria songarica)、盐爪爪(Kalidium foliatum)、白刺(Nitraria tangutorum)等]、山地旱生类群[狭叶锦鸡儿(Caragana stenophylla)、蒙古莸(Caryopteris mongholica)等]、山地中生类群[银露梅(Potentilla glabra)、金露梅(Potentilla fruticosa)、青海云杉(Picea crassifolia)等]、山地湿生类群[祁连圆柏(Sabina chinensis)、鬼箭锦鸡儿(Caragana jubata)、甘肃萱草(Carex kansuensis)等]和山地高寒类群[鬼箭锦鸡儿(Caragana jubata)、山生柳(Salix oritrepha)、青藏蓼(Koenigia fertilis Maxim)等]。
1.2 样方设置与调查
2017—2019年,以100 m海拔高程为间隔,在海拔1500~3700 m的范围内植被保存较好的区域设置30 m×30 m的样地92块,每块样地中随机布设15m×15m的乔木样方1个,在乔木样方中沿对角线随机布设灌木样方(5 m×5 m)和草本样方(1 m×1 m)各3个。样方调查中,详细记录个体的高度、多度、胸径、相对坐标等指标,同时记录样地的海拔、坡度、坡向、土壤性质等群落生境特征,气温和降水采用寺大隆气象站(海拔2 600 m)数据。共记录乔木7种、灌木14种、草本植物71种(表1)。
表 1 甘肃祁连山国家自然保护区植被优势种及重要值Table 1. Quantitative characteristics of dominant species in Qilian Mountain National Nature Reserve, Gansu Province种名 科名 属名 生活型 海拔范围/m 重要值 祁连圆柏 Sabina chinensis 柏科 圆柏属 乔木 3 050~3 300 201.43 青海云杉 Picea crassifolia 松科 云杉属 乔木 2 450~2 950 230.21 山杨 Populus daviviana 杨柳科 杨属 乔木 1 500~1 800 87.43 红桦 Betula albosinensis 桦木科 桦木属 乔木 2 450~2 700 90.12 白桦 Betula platyphylla 桦木科 桦木属 乔木 2 450~2 750 82.13 小叶杨 Populus simonii Carrière 杨柳科 杨属 乔木 1500~1700 40.21 狭叶锦鸡儿 Caragana stenophylla 豆科 锦鸡儿属 灌木 2300~2600 103.44 柽柳 Tamarix chinensis 柽柳科 柽柳属 灌木 1500~1700 43.23 鬼箭锦鸡儿 Caragana jubata 豆科 锦鸡儿属 灌木 3050~3700 110.76 山生柳 Salix oritrepha 杨柳科 柳属 灌木 3190~3700 56.27 金露梅 Potentilla fruticosa 蔷薇科 委陵菜属 灌木 2450~2950 98.51 银露梅 Potentilla glabra 蔷薇科 委陵菜属 灌木 2350~2600 72.61 白刺 Nitraria tangutorum 蒺藜科 白刺属 灌木 1500~1700 89.31 红砂 Reaumuria songarica 柽柳科 红砂属 灌木 1500~1800 80.10 盐爪爪 Kalidium foliatum 藜科 盐爪爪属 灌木 1500~1800 74.87 蒙古莸 Caryopteris mongholica 马鞭草科 莸属 灌木 2360~2750 65.32 甘蒙锦鸡儿 Caragana opulens 豆科 锦鸡儿属 灌木 2300~2360 55.34 合头草 Sympegma regelii 藜科 合头草属 灌木 1550~1730 67.89 冰草 Agropyron cristatum 禾本科 冰草属 多年生草本 1500~1700 95.38 芨芨草 Achnatherum splendens 禾本科 芨芨草属 多年生草本 1550~1730 89.38 青藏蓼 Koenigia fertilis Maxim 蓼科 冰岛蓼属 一年生草本 3190~3700 82.30 珠芽蓼 Bistorta vivipara (L.) Gray 蓼科 拳参属 多年生草本 3190~3700 89.50 牛筋草 Eleusine indica 禾本科 属 一年生草本 1500~1800 94.76 甘肃萱草 Carex kansuensis 莎草科 萱草属 多年生草本 2350~3300 96.31 甘青蒿 Artemisia tangutica 菊科 蒿属 多年生草本 3050~3300 90.17 垂穗披碱草 Elymus nutans 禾本科 披碱草属 多年生草本 2350~2950 89.88 藓生马先蒿 Pedicularis muscicola 玄参科 马先蒿属 多年生草本 2360~2750 40.08 1.3 数据处理与分析
分别计算每个样方群落:重要值(p)=(相对密度+相对频度+相对优势度)/3;Patrick指数R=S;D=1−
∑{Ni(Ni−1)/N(N−1)} ;Shannon-Wiener指数H=−∑(PilogPi) ;Pielou指数JSW =−(∑PilogPi)/logS [14]。其中,D表示优势度指数,N代表样地或层次的总重要值,Ni 代表第i个物种的重要值,Pi 为物种i的相对重要值,S为样方的物种总数。在上述统计的基础上,运用Cornel 3.5 完成样方物种信息的除趋势典范对应分析(DCCA),详细计算步骤参考Podobnik等[15]的研究报道,运用SPSS16.0完成样方物种信息与海拔梯度的相关分析及回归分析。为了更好地分析不同海拔植物群落多样性特征,本研究根据研究区生境和主要植被类型将海拔划分为低海拔(1 500~2 000 m)、中低海拔(2 000~2 400 m)、中海拔(2 400~2 950 m)和高海拔(2 950~3 700 m)4个梯度。2. 结果与分析
2.1 环境因子与DCCA排序轴的相关性
甘肃祁连山国家自然保护区植物群落DCCA排序结果见表2。由表2可以看出,草地群落前4个排序轴特征值分别为0.967,0.814,0.709和0.540,占总特征值的72.15%,对物种和环境关系的解释量累计为92.14%;灌木群落前4个排序轴的特征值分别为0.925,0.546,0.403和0.225,占总特征值的75.38%,对物种和环境关系的解释量累计为88.51%。
表 2 环境因子与DCCA前四排序轴的相关性Table 2. Correlation between environmental factors and the top four ranking axes of DCCA环境因子 草地层 灌木层 AX1 AX2 AX3 AX4 AX1 AX2 AX3 AX4 海拔 0.670** −0.125 −0.335* 0.103 0.786** −0.089 −0.085 0.121 年均温度 −0.592** 0.139 0.360* −0.092 −0.698** 0.124 0.150 −0.113 年均降雨 0.604** −0.135 −0.315* 0.092 0.726** −0.101 −0.079 0.058 坡度 0.584** −0.201 −0.259 0.067 0.429* −0.034 −0.501* 0.031 坡位 0.312* −0.184 0.209 −0.059 −0.486* −0.125 0.284 0.135 坡向 0.545** −0.317* 0.017 0.190 0.612** −0.238 0.241 −0.130 坡形 0.526** −0.246 0.015 0.014 0.120 0.015 −0.340 0.047 土壤腐殖质 0.523** −0.305 0.071 0.150 0.272 −0.102 0.216 −0.210 特征值 0.976 0.814 0.709 0.540 0.925 0.546 0.403 0.225 梯度长度 16.102 7.265 8.358 5.395 7.821 4.265 2.301 2.354 物种−环境关系的累计百分比方差 69.850 82.150 88.370 92.140 62.130 75.960 83.260 88.510 物种−环境因子相关性 0.968 0.845 0.678 0.621 0.921 0.832 0.625 0.619 注:*表示显著(P<0.05),**表示极显著(P<0.01)。 蒙特卡洛检验结果显示,各环境因子与草地群落和灌木群落前4个排序轴呈现出显著的相关性(P<0.01),其中第一轴蒙特卡洛检验结果在草地群落和灌木群落中分别为F=12.147,P=0.001和F=10.956,P=0.001。
草地群落排序中,各环境因子与DCCA第一轴均呈显著相关关系,相关系数大小表现为:海拔>年均降水>年均温度>坡度>坡向>坡形>坡位>土壤腐殖质。灌木群落DCCA第一轴所反映的环境梯度与草地类似,除坡形和土壤腐殖质外,各环境因子与第一轴均呈显著相关性,相关系数大小表现为:海拔>年均降雨>年均温度>坡向>坡位>坡度>土壤腐殖质>坡形。同时,DCCA第一轴在草地群落和灌木群落中均有最大的特征值和梯度长度,因此,样方DCCA第一轴排序位置代表了样方在环境空间中的生态位,通过各样方群落物种多样性的比较,可能会较客观地反映出多样性与环境梯度之间的内在联系[16]。
2.2 植物群落多样性在垂直环境梯度上的变化趋势
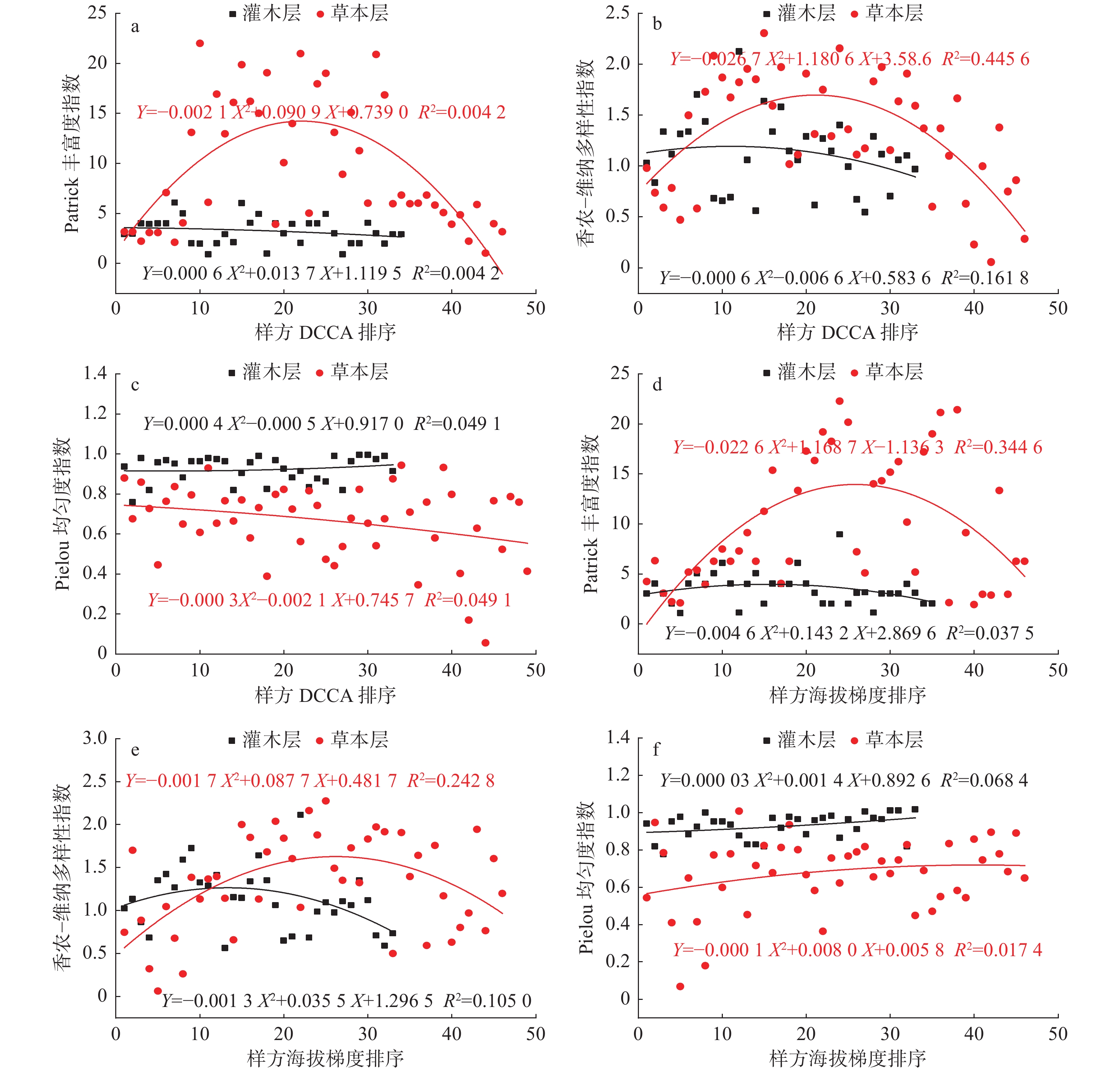
灌木群落和草本群落多样性在DCCA排序和海拔梯度排序上的分布格局见图2。由图2可以看出,甘肃祁连山国家自然保护区灌木层和草本层样方按DCCA第一轴排序和海拔梯度排序,Patrick丰富度指数和Shannon-Wiener多样性指数表现出较为一致的结果。灌木层样地间Patrick丰富度指数和Shannon-Wiener多样性指数在海拔梯度上有明显的波动,呈“单峰型”变化,但曲线拟合不显著(P>0.05)(图2 a、b);草本样地间Patrick丰富度指数和Shannon-Wiener多样性指数在海拔梯度和DCCA垂直环境梯度上均有强烈的波动(图2 a、b),呈显著(P<0.01)的“单峰型”变化,但相比海拔梯度排序,DCCA第一轴排序的峰值更靠右,多项式拟合优于灌木层。草本层和灌木层Pielou 均匀度指数虽有一定的波动,但整体的变化并不明显(P>0.05)(图2 a、b),说明植物群落的均匀度受环境梯度变化的影响较小。
![]() 图 2 灌木群落和草本群落多样性在DCCA排序和海拔梯度排序上的分布格局Figure 2. Distribution patterns of shrub and grassland community diversity in DCCA and altitudinal gradients
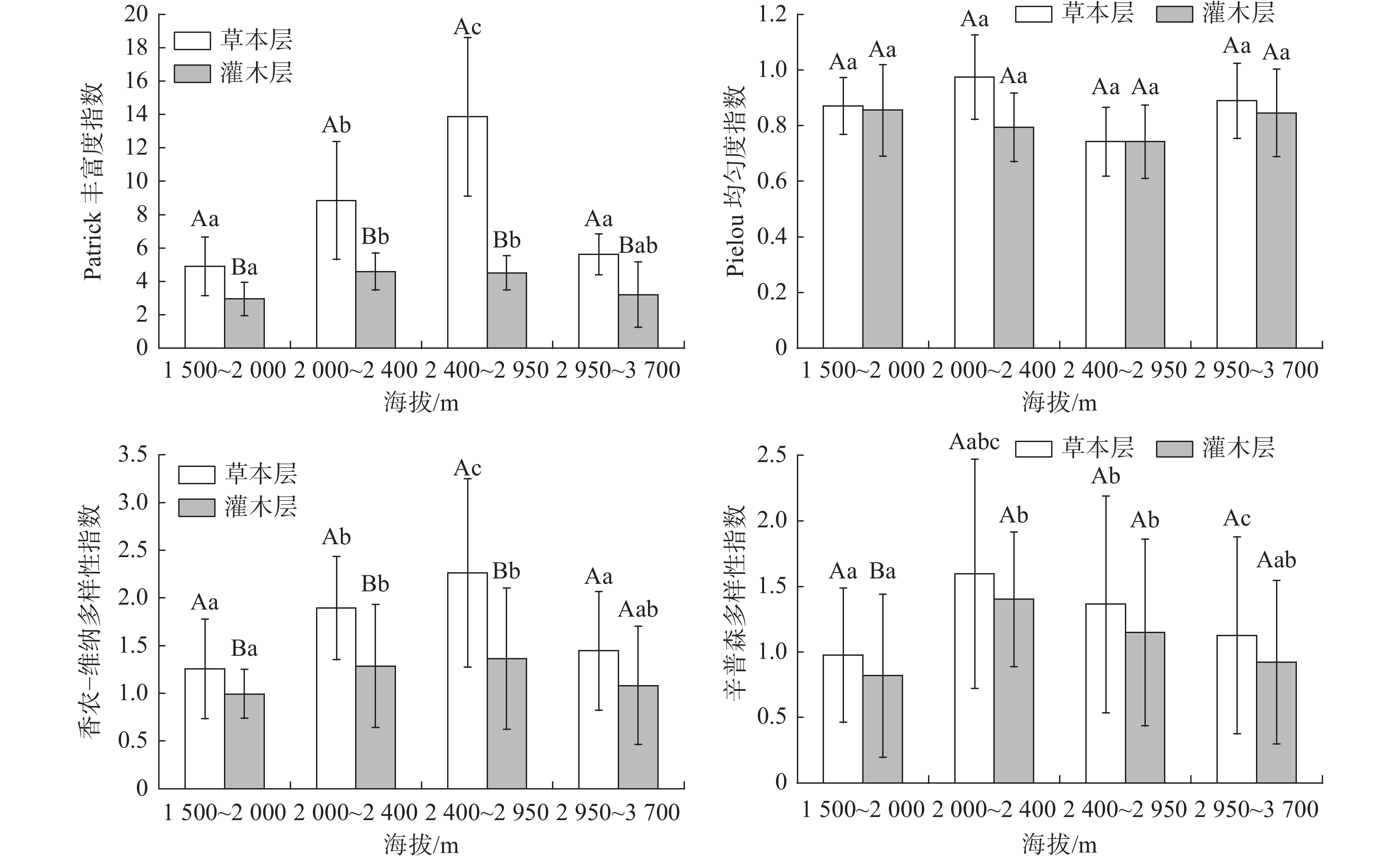
图 2 灌木群落和草本群落多样性在DCCA排序和海拔梯度排序上的分布格局Figure 2. Distribution patterns of shrub and grassland community diversity in DCCA and altitudinal gradients不同海拔草本层、灌木层植物群落多样性特征及差异显著性检验结果见图3。由图3可以看出,在不同海拔梯度植物群落丰富度指数、多样性指数和均匀度指数具有明显的差异:草本层Patrick丰富度指数、Shannon-Wiener多样性指数和Simpson多样性指数在低海拔地区(1 500~2 000 m)显著高于灌木层(P<0.01);在中低海拔地区(2 000~2 400 m)和中海拔地区(2 400~2 950 m),仅Patrick丰富度指数和Shannon-Wiener多样性指数显著高于灌木层(P<0.01),Simpson多样性指数无显著差异;在高海拔除Patrick丰富度指数达到显著相关外(P<0.05),Shannon-Wiener多样性指数和Simpson多样性指数均无显著差异,各植物群落均匀度在海拔梯度上未表现出差异性变化。差异显著性检验结果还表明,在中海拔地区(2 400~2 900 m):草本层的Patrick丰富度指数和Shannon-Wiener多样性指数最大,分别为13.867和2.264,均显著(P<0.01)高于低海拔和高海拔地区相应层的指数(图3);灌木层在中海拔地区(2400~2900 m)的Patrick丰富度指数和Shannon-Wiener多样性指数除与低海拔地区(1500~2000 m)有显著差异外(P<0.01),与其他海拔梯度均无显著差异。这与曲线的多项式拟合结果所反映的趋势较为一致(图1),说明海拔对草本层植物丰富度和多样性的影响较大。在样方调查中,共记录乔木7种,且分布较为均匀,其多样性指数在各环境梯度均未发现明显的差异,故未作比较。
![]() 图 3 不同海拔草本层、灌木层植物群落多样性特征及差异显著性检验注:图上不同大写字母表示同一海拔不同植被类型之间的差异;不同小写字母表示同一植被不同海拔高度之间的差异。Figure 3. Diversity characteristics and significance test of plant communities in herbaceous layer and shrub layer at different elevations
图 3 不同海拔草本层、灌木层植物群落多样性特征及差异显著性检验注:图上不同大写字母表示同一海拔不同植被类型之间的差异;不同小写字母表示同一植被不同海拔高度之间的差异。Figure 3. Diversity characteristics and significance test of plant communities in herbaceous layer and shrub layer at different elevations2.3 群落DCCA排序与海拔排序的关系
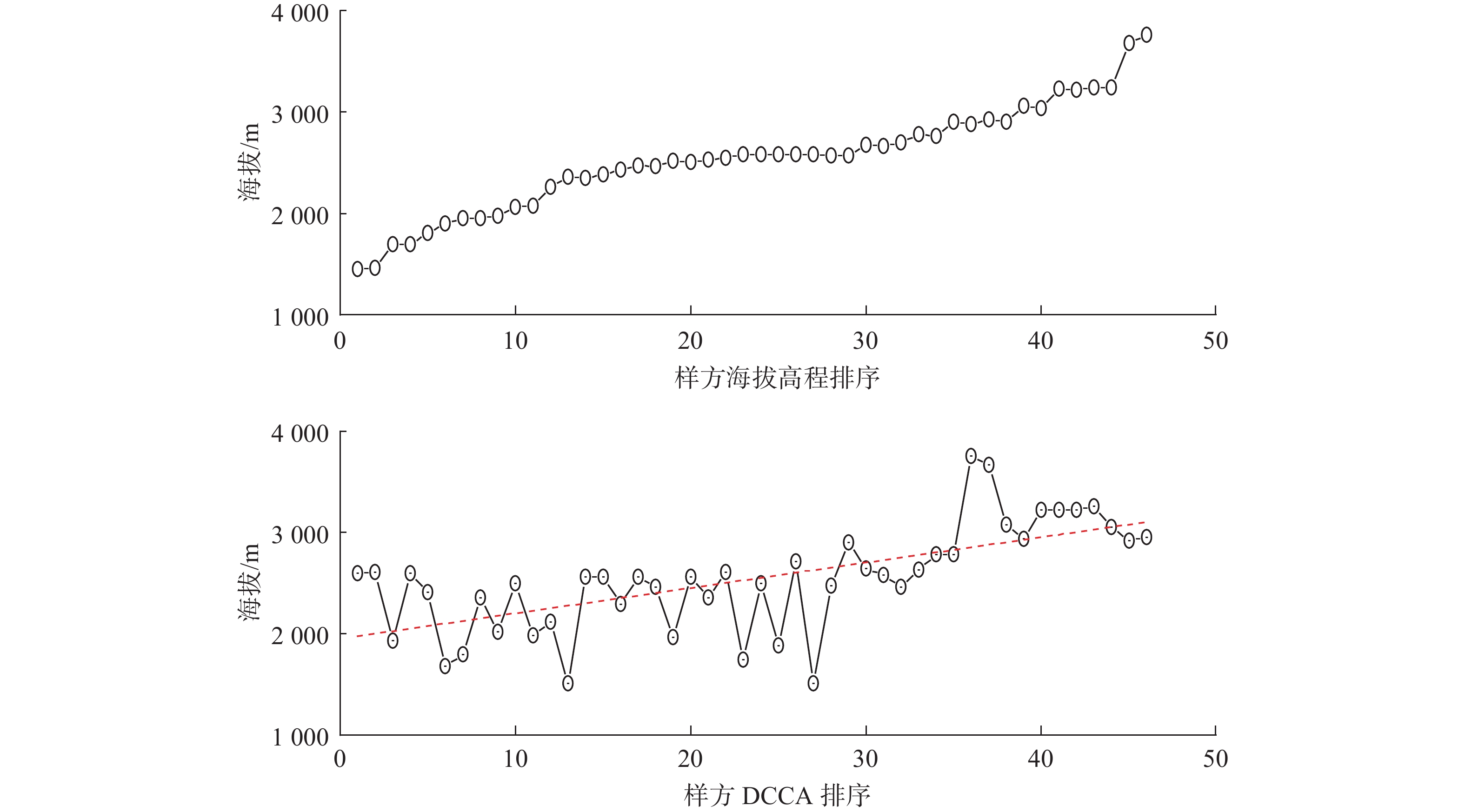
植物群落样方DCCA第一轴排列后样地海拔高程变化如图4所示。由图4可见,甘肃祁连山国家自然保护区草地群落样方DCCA排序与海拔高程排序表现出较为一致的变化趋势,各样方间海拔高程均由低到高变化,但海拔高程排序样方间海拔变化较为平缓,而DCCA排序样方间海拔高程具有剧烈的波动,说明DCCA排序对样方高程序列的变化具有显著的影响。植物群落分布格局与环境空间变量定量分析结果显示,环境因子对草本植物群落、灌木植物群落和乔木植物群落分布格局的解释量分别占29.32%,23.24%和9.05%,空间因子对草本植物群落、灌木植物群落和乔木植物群落分布格局的解释量分别占15.21%,11.08%和7.52%,空间和环境因子交叉作用对草本植物群落、灌木植物群落和乔木植物群落分布格局的解释量分别占11.04%,8.42%和5.31%,草本植物群落、灌木植物群落和乔木植物群落分布格局未被空间因子和环境因子解释部分分别占44.43%,57.26%和78.12%。
![]() 图 4 植物群落样方DCCA第一轴排列后样地海拔高程变化Figure 4. Elevation changes of the plant community quadrat after the first axis of DCCA arrangement
图 4 植物群落样方DCCA第一轴排列后样地海拔高程变化Figure 4. Elevation changes of the plant community quadrat after the first axis of DCCA arrangement3. 讨 论
植物多样性沿环境梯度的分布格局受气候、空间、环境等众多因子的影响[17]。本研究调查发现种子植物85种,隶属30科56属,禾本科、菊科和豆科的物种在研究区内占主要优势,这与唐志红等[18]对祁连山东段植物群落特征研究结果较为一致。研究区共发现乔木7种,主要以青海云杉为主,且有绝对优势,主要分布在中海拔的半阴坡和半阳坡,在高海拔的阳坡有少量祁连圆柏分布,在峡谷地带分布有少量的红桦和白桦,山杨在河谷地段成斑状分布。由于乔木层物种在甘肃祁连山国家自然保护区北坡分布较少,加之其个体生长缓慢,植被组成相对稳定,而灌木层和草本层植被物种多样性和丰富度却随着乔木层植被的演替而不断变化。灌木作为甘肃祁连山自然保护区北坡的主要植物类群,从低海拔到高海拔都有分布,共有13种,占植物种类的15.29%。此外,样方DCCA排序与海拔高程排序表现出较为一致的变化趋势,表明海拔作为影响山地植物分布的重要因素,在一定程度上代表着一个重要资源的生产力梯度[19]。但物种多样性沿海拔梯度的分布格局不仅与海拔选取范围有关,也应该注意土壤性质、人类活动、坡度坡向等其他因子对海拔引起的水热变化的再修饰[20]。甘肃祁连山国家自然保护区山体陡峭、地形复杂,海拔梯度效应在很大程度上被坡度、坡位等地形微环境削弱或加强[21]。由此可见,研究区海拔梯度并不能完全反映资源生产力梯度。DCCA作为植被环境关系多元分析强有力的工具,能够较为客观地反映样方资源环境生产力水平[22]。甘肃祁连山国家自然保护区样方DCCA排序后,物种多样性在样地间产生剧烈的波动,这与贺国宝等[23]研究结果较为一致,说明群落物种多样性除受海拔引起的水热梯度影响外,还与地形、人类活动、生物间相互作用等因素有密切的关系。
物种多样性受环境变化和人类活动的双重影响,是海拔引起的水热变化的综合体现,在一定程度上反应了群落的稳定性和复杂性[24]。祁连山国家自然保护区草地和灌木植物群落的Patrick丰富度指数和Shannon-Wiener多样性指数表现为“单峰型”曲线变化,在2 400~2 900 m的中海拔地带达到最高,这与钟娇娇等[25]在秦岭地区、刘彬等[26]等在天山南坡中段、贾文雄等[27]等在祁连山北坡植物群落特征研究中的结果相一致。在西北干旱半干旱地区,单峰型分布较为常见[28],由于研究区地处内陆,远离海洋,因而干燥少雨,年均降水量不足600 mm[29],加之区域内水土流失和荒漠化较为严重,物种多样性在很大程度上受降水和气温的影响[30]。在小尺度上,海拔变化致使浅山地带和高山地带分别受水分和气温的胁迫导致物种多样性较小[31],2 400~2 900 m的中海拔地带处于荒漠草原和草甸草原的过渡区,阳光充足、降水丰沛、气温适宜,土壤含水量升高,容重减小,孔隙度增加[32],有效地促进了土壤质地的改善和有机质含量的积累,增强了土壤持水保肥的能力。因此,在该地带发育了以冰草、赖草(Leymus secalinus)、针茅(Stipa capillata)为主的生态位较宽的禾本科植物群落和以甘蒙锦鸡儿、狭叶锦鸡儿为优势的喜光灌木群落,该地带植物群落种类繁多、结构复杂、生态位重叠,种间资源竞争激烈,伴生种在植物群落中占据了一定的生存空间,使得原有优势种逐渐弱化,物种的丰富度和多样性均较高[33]。高辉等[34]研究表明,在色季拉山,森林群落的均匀度在小尺度上除与海拔跨度相关外,样方面积也是影响其分布格局的重要因素,但在大尺度上植物群落的均匀度没有明显的变化。本研究还发现,样方DCCA排序和海拔高程排序,均匀度均未随环境梯度的变化而表现出特定的规律,由此说明植物群落均匀度受环境的影响较小,而主要受自身结构类型和稳定程度的影响[35]。
甘肃祁连山国家自然保护区物种多样性和丰富度垂直格局在不同层次群落之间虽表现出一定的共性,但在同一海拔范围内草本层的多样性和丰富度均显著(P<0.01)高于灌木层,该结论与李巧燕等[7]在长江三峡库区物种多样性垂直分布格局研究中的结果相一致。不同功能类型、不同进化历史的植物群落多样性和丰富度分布格局的差异,可能体现了特定植物群落对区域宏观气候的直接响应和对环境的生态适应性机制[36]。
4. 结 论
1)甘肃祁连山国家自然保护区草本层和灌木层Patrick丰富度指数和Shannon-Wiener多样性指数随环境梯度和海拔梯度均表现出“单峰型”变化趋势,但曲线拟合草本层优于灌木层,其变化敏感度依次表现为草本层>灌木层>乔木层;
2)甘肃祁连山国家自然保护区不同层次植物群落丰富度指数、多样性指数和均匀度指数在不同海拔表现出明显的差异,除高海拔(2 950~3 700 m)外,在其余海拔梯度上草本层丰富度指数和多样性指数均显著高于灌木层(P<0.01),且在中海拔(2 400~2 950 m)地带达到最高;
3)甘肃祁连山国家自然保护区植物群落不同层次均匀度在整个海拔梯度内均未表现出明显的差异,反映了植物群落均匀度受环境影响较小,而主要受自身结构类型和稳定程度的影响;
4)在影响植物群落空间分布格局的变量中,环境因子对草本、灌木和乔木植物群落分布格局的解释量分别占29.32%,23.24%和9.05%,空间因子的解释量分别占15.21%,11.08%和7.52%,空间和环境因子交叉作用的解释量分别占11.04%,8.42%和5.31%,未被空间因子和环境因子解释部分分别占44.43%,57.26%和78.12%。
-
![]()
图 2 灌木群落和草本群落多样性在DCCA排序和海拔梯度排序上的分布格局
Figure 2. Distribution patterns of shrub and grassland community diversity in DCCA and altitudinal gradients
![]()
图 3 不同海拔草本层、灌木层植物群落多样性特征及差异显著性检验
注:图上不同大写字母表示同一海拔不同植被类型之间的差异;不同小写字母表示同一植被不同海拔高度之间的差异。
Figure 3. Diversity characteristics and significance test of plant communities in herbaceous layer and shrub layer at different elevations
![]()
图 4 植物群落样方DCCA第一轴排列后样地海拔高程变化
Figure 4. Elevation changes of the plant community quadrat after the first axis of DCCA arrangement
表 1 甘肃祁连山国家自然保护区植被优势种及重要值
Table 1 Quantitative characteristics of dominant species in Qilian Mountain National Nature Reserve, Gansu Province
种名 科名 属名 生活型 海拔范围/m 重要值 祁连圆柏 Sabina chinensis 柏科 圆柏属 乔木 3 050~3 300 201.43 青海云杉 Picea crassifolia 松科 云杉属 乔木 2 450~2 950 230.21 山杨 Populus daviviana 杨柳科 杨属 乔木 1 500~1 800 87.43 红桦 Betula albosinensis 桦木科 桦木属 乔木 2 450~2 700 90.12 白桦 Betula platyphylla 桦木科 桦木属 乔木 2 450~2 750 82.13 小叶杨 Populus simonii Carrière 杨柳科 杨属 乔木 1500~1700 40.21 狭叶锦鸡儿 Caragana stenophylla 豆科 锦鸡儿属 灌木 2300~2600 103.44 柽柳 Tamarix chinensis 柽柳科 柽柳属 灌木 1500~1700 43.23 鬼箭锦鸡儿 Caragana jubata 豆科 锦鸡儿属 灌木 3050~3700 110.76 山生柳 Salix oritrepha 杨柳科 柳属 灌木 3190~3700 56.27 金露梅 Potentilla fruticosa 蔷薇科 委陵菜属 灌木 2450~2950 98.51 银露梅 Potentilla glabra 蔷薇科 委陵菜属 灌木 2350~2600 72.61 白刺 Nitraria tangutorum 蒺藜科 白刺属 灌木 1500~1700 89.31 红砂 Reaumuria songarica 柽柳科 红砂属 灌木 1500~1800 80.10 盐爪爪 Kalidium foliatum 藜科 盐爪爪属 灌木 1500~1800 74.87 蒙古莸 Caryopteris mongholica 马鞭草科 莸属 灌木 2360~2750 65.32 甘蒙锦鸡儿 Caragana opulens 豆科 锦鸡儿属 灌木 2300~2360 55.34 合头草 Sympegma regelii 藜科 合头草属 灌木 1550~1730 67.89 冰草 Agropyron cristatum 禾本科 冰草属 多年生草本 1500~1700 95.38 芨芨草 Achnatherum splendens 禾本科 芨芨草属 多年生草本 1550~1730 89.38 青藏蓼 Koenigia fertilis Maxim 蓼科 冰岛蓼属 一年生草本 3190~3700 82.30 珠芽蓼 Bistorta vivipara (L.) Gray 蓼科 拳参属 多年生草本 3190~3700 89.50 牛筋草 Eleusine indica 禾本科 属 一年生草本 1500~1800 94.76 甘肃萱草 Carex kansuensis 莎草科 萱草属 多年生草本 2350~3300 96.31 甘青蒿 Artemisia tangutica 菊科 蒿属 多年生草本 3050~3300 90.17 垂穗披碱草 Elymus nutans 禾本科 披碱草属 多年生草本 2350~2950 89.88 藓生马先蒿 Pedicularis muscicola 玄参科 马先蒿属 多年生草本 2360~2750 40.08  下载: 导出CSV
下载: 导出CSV
表 2 环境因子与DCCA前四排序轴的相关性
Table 2 Correlation between environmental factors and the top four ranking axes of DCCA
环境因子 草地层 灌木层 AX1 AX2 AX3 AX4 AX1 AX2 AX3 AX4 海拔 0.670** −0.125 −0.335* 0.103 0.786** −0.089 −0.085 0.121 年均温度 −0.592** 0.139 0.360* −0.092 −0.698** 0.124 0.150 −0.113 年均降雨 0.604** −0.135 −0.315* 0.092 0.726** −0.101 −0.079 0.058 坡度 0.584** −0.201 −0.259 0.067 0.429* −0.034 −0.501* 0.031 坡位 0.312* −0.184 0.209 −0.059 −0.486* −0.125 0.284 0.135 坡向 0.545** −0.317* 0.017 0.190 0.612** −0.238 0.241 −0.130 坡形 0.526** −0.246 0.015 0.014 0.120 0.015 −0.340 0.047 土壤腐殖质 0.523** −0.305 0.071 0.150 0.272 −0.102 0.216 −0.210 特征值 0.976 0.814 0.709 0.540 0.925 0.546 0.403 0.225 梯度长度 16.102 7.265 8.358 5.395 7.821 4.265 2.301 2.354 物种−环境关系的累计百分比方差 69.850 82.150 88.370 92.140 62.130 75.960 83.260 88.510 物种−环境因子相关性 0.968 0.845 0.678 0.621 0.921 0.832 0.625 0.619 注:*表示显著(P<0.05),**表示极显著(P<0.01)。
下载: 导出CSV
-
[1] 郑鸾, 龙翠玲. 茂兰喀斯特森林不同地形植物多样性与土壤理化特征研究[J]. 广西植物, 2020, 40(6): 792−801. DOI: 10.11931/guihaia.gxzw201906034. ZHENG L, LONG C L. Differences of plant diversity and soil physicochemical properties in Maolan Karst forest under different topographic conditions[J]. Guihaia, 2020, 40(6): 792−801. DOI: 10.11931/guihaia.gxzw201906034
[2] 刘秉儒. 生物多样性的海拔分布格局研究及进展[J]. 生态环境学报, 2021, 30(2): 438−444. DOI: 10.16258/j.cnki.1674-5906.2021.02.0025. LIU B R. Recent advances in altitudinal distribution patterns of biodiversity[J]. Ecology and Environmental Sciences, 2021, 30(2): 438−444. DOI: 10.16258/j.cnki.1674-5906.2021.02.0025
[3] 何远政, 黄文达, 赵昕, 等. 气候变化对植物多样性的影响研究综述[J]. 中国沙漠, 2021, 41(1): 59−66. DOI: 10.7522/j.issn.1000-694X.2020.00104. HE Y Z, HUANG W D, ZHAO X, et al. Review on the impact of climate change on plant diversity[J]. Journal of Desert Research, 2021, 41(1): 59−66. DOI: 10.7522/j.issn.1000-694X.2020.00104
[4] 陈水飞, 徐辉, 林文俊, 等. 武夷山国家公园植物群落物种多样性沿海拔梯度的变化分析[J]. 植物资源与环境学报, 2023, 32(1): 1−9. DOI: 10.3969/j.issn.1674-7895.2023.01.01. CHEN S F, XU H, LIN W J, et al. Variation analysis on species diversity of plant communities along the elevation gradient in Wuyishan National Park[J]. Journal of Plant Resources and Environment, 2023, 32(1): 1−9. DOI: 10.3969/j.issn.1674-7895.2023.01.01
[5] 王褚, 王世伟, 潘存德, 等. 天山北坡中部天山云杉林植物多样性沿海拔变化特征[J]. 新疆农业大学学报, 2021, 44(3): 172−178. DOI: 10.3969/j.issn.1007-8614.2021.03.003. WANG C, WANG S W, PAN C D, et al. Variation characteristics of plant diversity along altitude in Picea schrenk-iana forestin the central part of the northern slope of Tianshan Mountains[J]. Journal of Xinjiang Agricultural University, 2021, 44(3): 172−178. DOI: 10.3969/j.issn.1007-8614.2021.03.003
[6] 徐翔, 张化永, 谢婷, 等. 西双版纳种子植物物种多样性的垂直格局及机制[J]. 生物多样性, 2018, 26(7): 678−689. DOI: 10.17520/biods.2017132. XU X, ZHANG H Y, XIE T, et al. Elevational pattern of seed plant diversity in Xishuangbanna and its mechanisms[J]. Biodiversity Science, 2018, 26(7): 678−689. DOI: 10.17520/biods.2017132
[7] 李巧燕, 王襄平. 长江三峡库区物种多样性的垂直分布格局: 气候、几何限制、面积及地形异质性的影响[J]. 生物多样性, 2013, 21(2): 141−152. DOI: 10.3724/SP.J.1003.2013.12183. LI Q Y, WANG X P. Elevational pattern of species richness in the Three Gorges region of the Yangtze River: effect of climate, geometric constraints, area and topographical heterogeneity[J]. Biodiversity Science, 2013, 21(2): 141−152. DOI: 10.3724/SP.J.1003.2013.12183
[8] 董晓红. 祁连山排露沟小流域森林植被水文影响的模拟研究[D]. 北京: 中国林业科学研究院, 2007. DONG X H. A study on modelling the hydrological effect of forest and vegetation in the pailugou catchment, Qilian Mountains[D]. Beijing: Chinese Academy of Forestry, 2007.
[9] 杨荣荣, 曹广超, 曹生奎, 等. 祁连山南坡主要河谷NDVI时空变化及影响因素分析[J]. 广西植物, 2021, 41(3): 429−437. DOI: 10.11931/guihaia.gxzw201907036. YANG R R, CAO G C, CAO S K, et al. Temporal and spatial variations of NDVI and analysis of influencing factors in main valleys of southern slope of Qilian Mountains[J]. Guihaia, 2021, 41(3): 429−437. DOI: 10.11931/guihaia.gxzw201907036
[10] ZHU H, CAO K, CHEN X L, et al. Effects of suspended ecological beds on phytoplankton community structure in Baiyangdian Lake, China[J]. Journal of Freshwater Ecology, 2022, 37(1): 189−202. DOI: 10.1080/02705060.2022.2036646.
[11] 刘菊梅. 乌梁素海湿地挺水植物根圈脱氮甲烷氧化菌群多样性及分布特征研究[D]. 呼和浩特: 内蒙古大学, 2018. LIU J M. Diversity of denitrifying methantrophs and their distribution in root zone of three emergent macrophytes in eutrophic Wuliangsuhai wetland[D]. Hohhot: Inner Mongolia University, 2018.
[12] 李玉辰, 李宗省, 张小平, 等. 祁连山国家公园植被时空变化及其对人类活动的响应[J]. 生态学报, 2023, 43(1): 219−233. DOI: 10.5846/stxb202107221990. LI Y C, LI Z X, ZHANG X P, et al. Spatial and temporal changes in vegetation cover and response to human activities in Qilian Mountain National Park[J]. Acta Ecologica Sinica, 2023, 43(1): 219−233. DOI: 10.5846/stxb202107221990
[13] 苏军德, 蒲济林, 李国霞, 等. 甘肃祁连山国家自然保护区生态系统效益评价研究[J]. 生态科学, 2021, 40(5): 89−94. DOI: 10.14108/j.cnki.1008-8873.2021.05.011. SU J D, PU J L, LI G X, et al. Study on ecosystem benefit evaluation of Qilian Mountains National Nature Reserve of Gansu Province[J]. Ecological Science, 2021, 40(5): 89−94. DOI: 10.14108/j.cnki.1008-8873.2021.05.011
[14] 周康. 海南热带雨林国家公园兰科植物多样性及其优先保护策略[D]. 海口: 海南大学, 2021. ZHOU K. Diversity of orchid in Hainan Tropical Rainforest National Park and its priority protection strategy[D]. Haikou: Hainan University, 2021.
[15] PODOBNIK B, STANLEY H E. Detrended cross-correlation analysis: a new method for analyzing two nonstationary time series[J]. Physical Review Letters, 2008, 100(8): 084102. DOI: 10.1103/physrevlett.100.084102.
[16] ZHANG Z, CHEN H B, HUANG X L. Prediction of air quality combining wavelet transform, DCCA correlation analysis and LSTM model[J]. Applied Sciences, 2023, 13(5): 2796. DOI: 10.3390/app13052796.
[17] ZHENG Z Q, CHEN T T. Study on distribution pattern of plant diversity of forest vegetation in luofu mountain, Guangdong[J]. Chinese Forestry Science and Technology, 2007, 6(2): 83−92.
[18] 唐志红, 尉秋实, 刘虎俊, 等. 祁连山东段高寒植被群落特征及其与地形气候因子关系研究[J]. 生态学报, 2020, 40(1): 223−232. DOI: 10.5846/stxb201812192760. TANG Z H, YU Q S, LIU H J, et al. Characteristics of alpine vegetation community and its relationship to topographic climate factors in the eastern Qilian Mountain[J]. Acta Ecologica Sinica, 2020, 40(1): 223−232. DOI: 10.5846/stxb201812192760
[19] 姜祖扬, 覃业辉, 刘季, 等. 海拔和林分密度对热带雨林林下植物多样性的影响[J]. 生态学杂志, 2023, 42(5): 1049−1055. DOI: 10.13292/j.1000-4890.202305.023. JIANG Z Y, QIN Y H, LIU J, et al. Effects of altitude and stand density on understory plant diversity in tropical rainforests[J]. Chinese Journal of Ecology, 2023, 42(5): 1049−1055. DOI: 10.13292/j.1000-4890.202305.023
[20] 木拉提江·夏地买买提, 艾克拜尔·伊拉洪, 阿马努拉·依明尼亚孜. 坡向和海拔对乌孙山草地植物群落特征及物种多样性的影响[J]. 新疆农业科学, 2022, 59(11): 2724−2733. DOI: 10.6048/j.issn.1001-4330.2022.11.016. MULATIJIANG X, AIKEBAIER Y, AMANULA Y, et al. Effects of slope aspect and altitude on characteristics and diversity of grassland plant communities in Wusun mountain[J]. Xinjiang Agricultural Sciences, 2022, 59(11): 2724−2733. DOI: 10.6048/j.issn.1001-4330.2022.11.016
[21] 苏军德, 赵晓冏, 李国霞. 甘肃祁连山国家自然保护区植物群落分布格局及其与环境因子的关系[J]. 广西植物, 2022, 42(3): 450−459. DOI: 10.11931/guihaia.gxzw202101034. SU J D, ZHAO X J, LI G X. Distribution pattern of plant community and its relationship with environmental factors in Qilian Mountains National Nature Reserve of Gansu Province[J]. Guihaia, 2022, 42(3): 450−459. DOI: 10.11931/guihaia.gxzw202101034
[22] 张雨桐, 刘华, 陈永富, 等. 大岗山植物群落物种多样性与群落结构研究[J]. 热带作物学报, 2023, 44(5): 1052−1061. DOI: 10.3969/j.issn.1000-2561.2023.05.020. ZHANG Y T, LIU H, CHEN Y F, et al. Species diversity and structure of tree population in Dagangshan[J]. Chinese Journal of Tropical Crops, 2023, 44(5): 1052−1061. DOI: 10.3969/j.issn.1000-2561.2023.05.020
[23] 贺国宝. 祁连山北坡植物群落空间分布格局与多样性研究[J]. 草业学报, 2021, 30(12): 194−201. DOI: 10.11686/cyxb2020463. HE G B. Distribution characteristics and plant community diversity on the north slopes of the Qilian Mountains[J]. Acta Prataculturae Sinica, 2021, 30(12): 194−201. DOI: 10.11686/cyxb2020463
[24] 张沛健, 徐建民, 卢万鸿, 等. 雷州半岛不同林龄尾细桉人工林植物多样性和土壤理化性质分析[J]. 中南林业科技大学学报, 2021, 41(9): 96−105. DOI: 10.14067/j.cnki.1673-923x.2021.09.011. ZHANG P J, XU J M, LU W H, et al. Plant diversity and soil physicochemical properties under different aged Eucalyptus urophylla × Eucalyptus tereticornis plantations in Leizhou Peninsula[J]. Journal of Central South University of Forestry & Technology, 2021, 41(9): 96−105. DOI: 10.14067/j.cnki.1673-923x.2021.09.011
[25] 钟娇娇, 陈杰, 陈倩, 等. 秦岭山地天然次生林群落MRT数量分类、CCA排序及多样性垂直格局[J]. 生态学报, 2019, 39(1): 277−285. DOI: 10.5846/stxb201712282338. ZHONG J J, CHEN J, CHEN Q, et al. Quantitative classification of MRT, CCA ordination, and species diversity along elevation gradients of a natural secondary forest in the Qinling Mountains[J]. Acta Ecologica Sinica, 2019, 39(1): 277−285. DOI: 10.5846/stxb201712282338
[26] 刘彬, 布买丽娅木·吐如汗, 艾比拜姆·克热木, 等. 新疆天山南坡中段种子植物区系垂直分布格局分析[J]. 植物科学学报, 2018, 36(2): 191−202. DOI: 10.11913/PSJ.2095-0837.2018.20191. LIU B, BUMARYAM T, AIBIBAIMU K, et al. Vertical distribution patterns of the seed plant flora in the middle section on the southern slopes of the Tianshan Mountains in Xinjiang, China[J]. Plant Science Journal, 2018, 36(2): 191−202. DOI: 10.11913/PSJ.2095-0837.2018.20191
[27] 贾文雄, 陈京华, 张禹舜, 等. 祁连山北坡草地植物群落特征与土壤水热因子的关系[J]. 生态学杂志, 2016, 35(3): 661−667. DOI: 10.13292/j.1000-4890.201603.032. JIA W X, CHEN J H, ZHANG Y S, et al. The relationship of characteristics of meadow communities with soil moisture and temperature in the northern slope of Qilian Mountains[J]. Chinese Journal of Ecology, 2016, 35(3): 661−667. DOI: 10.13292/j.1000-4890.201603.032
[28] 李红霖, 贺丽, 吴科君, 等. 黄河上游白河干流全段植物群落特征及生物多样性[J]. 草业科学, 2023, 40(4): 848−863. DOI: 10.11829/j.issn.1001-0629.2022-0303. LI H L, HE L, WU K J, et al. Research on plant community characteristics and biodiversity in the whole section of the main stream of Baihe River in the upper reaches of the Yellow River[J]. Pratacultural Science, 2023, 40(4): 848−863. DOI: 10.11829/j.issn.1001-0629.2022-0303
[29] 任婧, 张文煜, 黄颖, 等. 祁连山区再分析降水数据的适用性分析[J]. 兰州大学学报(自然科学版), 2023, 59(1): 23−28. DOI: 10.13885/j.issn.0455-2059.2023.01.004. REN J, ZHANG W Y, HUANG Y, et al. Applicability analysis of reanalysis precipitation data in the Qilian Mountains[J]. Journal of Lanzhou University (Natural Sciences), 2023, 59(1): 23−28. DOI: 10.13885/j.issn.0455-2059.2023.01.004
[30] 王有恒, 李丹华, 卢国阳, 等. 祁连山气候变化特征及其对水资源的影响[J]. 应用生态学报, 2022, 33(10): 2805−2812. DOI: 10.13287/j.1001-9332.202210.024. WANG Y H, LI D H, LU G Y, et al. Characteristics of climate change and its impacts on water resources in Qilian Mountains, China[J]. Chinese Journal of Applied Ecology, 2022, 33(10): 2805−2812. DOI: 10.13287/j.1001-9332.202210.024
[31] 余建平, 陈小南, 陈声文. 钱江源国家公园生物多样性监测体系研究[J]. 自然保护地, 2021, 1(2): 58−67. DOI: 10.12335/2096-8981.2020121401. YU J P, CHEN X N, CHEN S W. Study on biodiversity monitoring system of Qianjiangyuan National Park[J]. Natural Protected Areas, 2021, 1(2): 58−67. DOI: 10.12335/2096-8981.2020121401
[32] 于水今, 王娟, 何海燕, 等. 针阔混交林生物量稳定性驱动因子[J]. 林业科学, 2022, 58(11): 181−190. DOI: 10.11707/j.1001-7488.20221117. YU S J, WANG J, HE H Y, et al. Driving factors of the temporal stability of biomass of mixed broadleaf-conifer forest[J]. Scientia Silvae Sinicae, 2022, 58(11): 181−190. DOI: 10.11707/j.1001-7488.20221117
[33] 马桂英. 祁连山六大流域浮游生物群落结构特征及其影响因素研究[D]. 兰州: 兰州大学, 2022: 85-90. MA G Y. The characteristics and influencing factors of plankton community structure in six basins of Qilian Mountains[D]. Lanzhou: Lanzhou University, 2022: 85-90.
[34] 高辉, 刘丽娟, 方江平. 色季拉山沿海拔梯度的森林群落分布格局及环境解释[J]. 湖南师范大学自然科学学报, 2020, 43(5): 39−47,64. DOI: 10.7612/j.issn.2096-5281.2020.05.006. GAO H, LIU L J, FANG J P. Distribution pattern and environmental interpretation of forest communities along elevation gradients in Sejila Mountain[J]. Journal of Natural Science of Hunan Normal University, 2020, 43(5): 39−47,64. DOI: 10.7612/j.issn.2096-5281.2020.05.006
[35] 刘永靖. 祁连山中东部草本植物群落谱系结构特征及其影响因素[D]. 兰州: 兰州大学, 2021. LIU Y J. Characteristics and influencing factors of the phylogenetic structure of herbaceous plant communities in the middle-eastern Qilian Mountains[D]. Lanzhou: Lanzhou University, 2021.
[36] 袁荣珍, 王国宏, 唐志尧, 等. 红皮云杉林的植被分类及其环境解释[J]. 植物学报, 2022, 57(4): 468−478. DOI: 10.11983/CBB21181. YUAN R Z, WANG G H, TANG Z Y, et al. The vegetation classification on Picea koraiensis forest alliance and its environmental interpretation[J]. Chinese Bulletin of Botany, 2022, 57(4): 468−478. DOI: 10.11983/CBB21181
-
期刊类型引用(5)
1. 龚昕,霍新茹,李雯,杨彦东,刘超,秦伟春,沈艳,王国会,马红彬. 宁夏罗山山地草原植被群落特征及其空间分异. 草业学报. 2025(02): 1-15 .  百度学术
百度学术
2. 刘悦,徐杰. 青海省境内祁连山苔藓植物多样性及区系特征. 中国野生植物资源. 2025(02): 101-107 . 百度学术
3. 王润,徐明,杨雪,田粤,张健. 黔中地区不同马尾松针阔混交林群落结构、多样性及空间分布特征. 山地农业生物学报. 2025(02): 87-92 . 百度学术
4. 杨燕,张晓荣,刘晓东. 乌兰察布市公园植物多样性保护策略探析. 乡村科技. 2025(07): 111-115 . 百度学术
5. 莫舜华,胡兴华,叶家桐,周杨名. 银竹老山杜鹃花群落物种多样性沿海拔梯度分布格局. 中国野生植物资源. 2025(04): 113-119 . 百度学术
其他类型引用(6)






计量
- 文章访问数: 161
- HTML全文浏览量: 46
- PDF下载量: 67
- 被引次数: 11

 邮件订阅
邮件订阅 Rss
Rss